LES PROTÉINES ET L'OUBLI
Des
neuroscientifiques de l’Ecole polytechnique fédérale de Zurich (EPFZ) se sont
intéressés au rôle d’une protéine dans l’oubli, la PP1, déjà connue pour être
impliquée dans les mécanismes de l’apprentissage. Pour étudier la fonction de
cette protéine sur l’oubli, les chercheurs ont modifié génétiquement des
groupes de souris afin de pouvoir facilement activer et désactiver la protéine.
Puis ils ont effectué une série d’expériences sur la mémorisation et la
persistance des souvenirs.
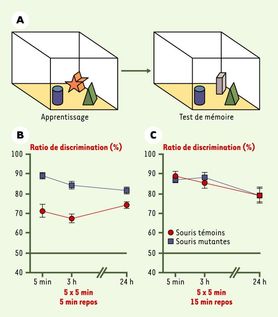
La première expérience a consisté à tester la mémorisation chez un groupe de souris témoins, et chez un groupe de souris mutantes chez lesquelles l’activité de la protéine PP1 a été désactivée. L’exercice de mémorisation consiste à tout d’abord faire mémoriser la position de trois objets dans une pièce par chaque souris. Les souris ont 5x5 minutes d’entraînement espacées soit de 5 minutes de pause, soit de 15 minutes de pause. Puis, l’un des objets est remplacé par un autre objet. Le groupe de souris s’attardant le plus sur le nouvel objet sera le groupe chez lequel la mémorisation a été la plus efficace. Les résultats sont nets : lorsqu’il y a seulement 5 minutes de pause, le ratio de discrimination (pourcentage de temps passé à s’attarder sur le nouvel objet) est plus élevé chez les souris mutantes, que ce soit, 5 minutes, 3 heures ou même 24 heures après l’expérience. Pour réussir à égaler les capacités de mémorisation des souris mutantes, les souris témoins doivent bénéficier de longues pauses entre chaque entraînement.
La protéine PP1 agit donc au cours de la mémorisation en freinant la formation du souvenir. Cependant, de longues pauses entre les séances d’apprentissage permettent de contrecarrer cet effet.
La première expérience a consisté à tester la mémorisation chez un groupe de souris témoins, et chez un groupe de souris mutantes chez lesquelles l’activité de la protéine PP1 a été désactivée. L’exercice de mémorisation consiste à tout d’abord faire mémoriser la position de trois objets dans une pièce par chaque souris. Les souris ont 5x5 minutes d’entraînement espacées soit de 5 minutes de pause, soit de 15 minutes de pause. Puis, l’un des objets est remplacé par un autre objet. Le groupe de souris s’attardant le plus sur le nouvel objet sera le groupe chez lequel la mémorisation a été la plus efficace. Les résultats sont nets : lorsqu’il y a seulement 5 minutes de pause, le ratio de discrimination (pourcentage de temps passé à s’attarder sur le nouvel objet) est plus élevé chez les souris mutantes, que ce soit, 5 minutes, 3 heures ou même 24 heures après l’expérience. Pour réussir à égaler les capacités de mémorisation des souris mutantes, les souris témoins doivent bénéficier de longues pauses entre chaque entraînement.
La protéine PP1 agit donc au cours de la mémorisation en freinant la formation du souvenir. Cependant, de longues pauses entre les séances d’apprentissage permettent de contrecarrer cet effet.
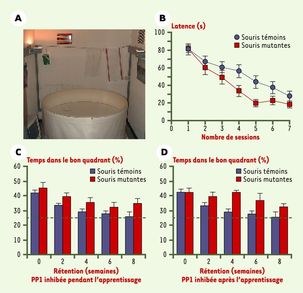
La deuxième expérience avait pour but de déterminer le rôle de la PP1 dans la persistance d’un souvenir dans le temps chez trois groupes de souris : un groupe témoin, un dont la PP1 a été désactivée uniquement pendant l’apprentissage, et un autre dont la PP1 a été désactivée après l’apprentissage. Les trois groupes ont mémorisé l’emplacement d’une plateforme dans un bassin à l’aide d’un entraînement intensif. Les chercheurs ont déjà pu observer que les souris dont la PP1 a été désactivée pour l’apprentissage apprenaient plus vite que les autres. On a ensuite enlevé la plateforme et observé le pourcentage de temps que passaient les souris dans la partie où s’était trouvée la plateforme. L’expérience est réalisée le jour d’après, puis elle est répétée toutes les deux semaines pendant 8 semaines. On remarque que les souris mutantes passent plus de temps dans la bonne partie du bassin que les souris témoins, que ce soit juste après l’apprentissage ou 8 semaines après. Cependant, lorsque la PP1 est désactivée après l’apprentissage, le temps passé dans la bonne partie est supérieure à quand la protéine est seulement désactivée lors de l’apprentissage.
La PP1 a non seulement un impact négatif sur la mémorisation, mais également lors de la consolidation et du rappel d’un souvenir.
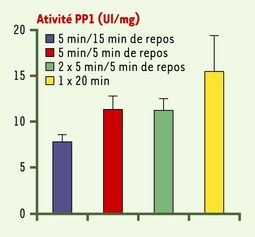
L’activité de la PP1 a été mesurée dans le cerveau de souris témoins lors de la première expérience, après :
- une session de 5 minutes suivie de 15 minutes de repos ;
- une session de 5 minutes suivie de 5 minutes de repos ;
- deux sessions de 5 minutes avec 5 minutes de pause entre les deux sessions, et 5 minutes de repos après les deux ;
- une session de 20 minutes sans repos.
On peut remarquer que plus le temps d’apprentissage est long et les pauses courtes, plus l’activité de la protéine est élevée. Ces résultats confirment ceux de la première expérience, comme quoi des pauses entre les sessions d’apprentissage permettent de faire baisser l’activité de PP1, et ainsi optimiser l’apprentissage.
- une session de 5 minutes suivie de 15 minutes de repos ;
- une session de 5 minutes suivie de 5 minutes de repos ;
- deux sessions de 5 minutes avec 5 minutes de pause entre les deux sessions, et 5 minutes de repos après les deux ;
- une session de 20 minutes sans repos.
On peut remarquer que plus le temps d’apprentissage est long et les pauses courtes, plus l’activité de la protéine est élevée. Ces résultats confirment ceux de la première expérience, comme quoi des pauses entre les sessions d’apprentissage permettent de faire baisser l’activité de PP1, et ainsi optimiser l’apprentissage.
A présent, nous allons expliquer les phénomènes observés durant ces expériences. Nous avons pu voir dans la première partie sur le fonctionnement de la mémoire, que cette dernière est basée sur des protéines, comme la protéine CREB, qui permettent le renforcement des synapses, et ainsi, la potentialisation à long terme. Nous avons également vu que ces protéines étaient activées grâce à l’action des protéines kinases, des enzymes qui permettent l’activation de ces protéines de la mémoire. Et bien la PP1 tient le rôle exactement inverse de celui des protéines kinases. La PP1 fait partie de la famille des phosphatases, tout comme la calcineurine (PP2B) dont les résultats aux expériences précédentes sont sensiblement les mêmes que ceux de la PP1. Ce sont également des protéines enzymatiques qui agissent sur les protéines de la mémoire en les déphosphorylant, c’est-à-dire en leur ôtant un atome de phosphore, ce qui a pour effet de les désactiver, les empêchant ainsi de renforcer les liaisons synaptiques en créant de nouveaux neurorécepteurs. Cependant, leur action peut être déjouée en faisant des pauses régulières lors d’un apprentissage, comme tous les étudiants ont dû s’en rendre compte, car durant les pauses, les protéines kinases ont le temps de rephosphoryler les protéines de la mémoire. Malheureusement, en ce qui concerne les souvenirs à proprement parler, ceux de la mémoire épisodique, cette solution n’est pas envisageable, car les souvenirs autobiographiques se forment en permanence. Les protéines phosphatases ont non seulement un rôle de régulateurs lors de la mémorisation, mais également dans le maintien de souvenirs.
"La dépression à long terme [...] a pour but de faire un premier tri dans les souvenirs accumulés au cours de la journée."

En effet, les phosphatases agissent sur les neurones après la mémorisation, il s’agit de la dépression à long terme. La dépression à long terme est le phénomène inverse de celui de la potentialisation à long terme, vu dans la partie sur la consolidation des souvenirs. La dépression à long terme a lieu durant le sommeil, comme la potentialisation à long terme, mais pas durant la même phase. La dépression à long terme a lieu durant le sommeil lent, qui précède le sommeil paradoxal durant lequel a lieu la consolidation des souvenirs. Ce phénomène a pour but de faire un tri dans les souvenirs accumulés pendant la journée. Afin d’effectuer ce tri, ce serait l’hippocampe, qui, lors de l’encodage, marquerait les synapses des neurones formant les souvenirs considérés comme utiles. Cet « étiquetage » se produit grâce au circuit de Papez, et en particulier l’amygdale qui gère les émotions liées au souvenir. Lorsque l’encodage d’un souvenir correspond à une forte émotion, l’amygdale s’active plus fortement et entraîne le marquage des neurones de la nouvelle trace mnésique fraîchement formée. Ainsi, les synapses non étiquetées sont altérées par les protéines phosphatases. Une fois ce tri effectué, les souvenirs restants sont consolidés par la potentialisation à long terme. On ne sait pas encore comment l’hippocampe fait pour étiqueter les neurones, ni quelle est la nature de ces étiquettes.
L'oubli lié à la vieillesse
Une étude publiée par des chercheurs de l'université de Columbia de New York en 2013 a mis en évidence le rôle important d’une autre protéine, mais cette fois dans la déficience de la mémoire liée l’âge. Il s’agit de la protéine RbAp48, codée par le gène du même nom. Pour aboutir à ce résultat, les scientifiques ont étudié les neurones de personnes décédées à différents âges, de 33 à 88 ans. En réalisant une analyse génétique des cellules du gyrus dentelé, une région de l’hippocampe, ils se sont rendu compte que le gène RbAp48 était moins présent chez les sujets les plus âgés. Pour confirmer ces résultats, les chercheurs ont effectués des expériences sur des souris. Ils ont remarqué que, lorsque l’on désactive le gène chez de jeunes souris, ces dernières présentaient les mêmes troubles de la mémoire que des souris âgées. Mieux, les chercheurs ont augmenté l’activation du gène chez les vieilles souris, qui ont retrouvé une mémoire « normale ».
Ainsi, la perte de la mémoire due à la vieillesse n’est pas un premier symptôme de la maladie d’Alzheimer, comme le pensaient les scientifiques jusqu’alors. En effet, cette étude a montré que la perte de le mémoire liée à l’âge était un phénomène réversible, contrairement à la maladie d’Alzheimer, que nous allons étudier plus tard, et qui résulte, elle, de la destruction de neurones.
L'oubli lié à la vieillesse
Une étude publiée par des chercheurs de l'université de Columbia de New York en 2013 a mis en évidence le rôle important d’une autre protéine, mais cette fois dans la déficience de la mémoire liée l’âge. Il s’agit de la protéine RbAp48, codée par le gène du même nom. Pour aboutir à ce résultat, les scientifiques ont étudié les neurones de personnes décédées à différents âges, de 33 à 88 ans. En réalisant une analyse génétique des cellules du gyrus dentelé, une région de l’hippocampe, ils se sont rendu compte que le gène RbAp48 était moins présent chez les sujets les plus âgés. Pour confirmer ces résultats, les chercheurs ont effectués des expériences sur des souris. Ils ont remarqué que, lorsque l’on désactive le gène chez de jeunes souris, ces dernières présentaient les mêmes troubles de la mémoire que des souris âgées. Mieux, les chercheurs ont augmenté l’activation du gène chez les vieilles souris, qui ont retrouvé une mémoire « normale ».
Ainsi, la perte de la mémoire due à la vieillesse n’est pas un premier symptôme de la maladie d’Alzheimer, comme le pensaient les scientifiques jusqu’alors. En effet, cette étude a montré que la perte de le mémoire liée à l’âge était un phénomène réversible, contrairement à la maladie d’Alzheimer, que nous allons étudier plus tard, et qui résulte, elle, de la destruction de neurones.
